
Mikrobióm – „kompendium“ pre diabetológov
Autoři:
Ľubomír Skladaný 1,2; Pavel Kohout 3
Působiště autorů:
II. interná klinika SZU, FNsP F. D. Roosevelta, Banská Bystrica
1; II. interná klinika UNLP a LF UPJŠ Košice
2; Interní klinika 3. LF UK a Fakultní Thomayerova nemocnice, Praha
3
Vyšlo v časopise:
Forum Diab 2026; 15(1): 36-41
Kategorie:
Hlavná téma
Mikrobióm – jeden z najdôležitejších orgánov ľudského tela
„Je to jednoduché: mikróby vládnu svetu.“
Keď Joshua Lederberg v roku 2001 zaviedol termín mikrobióm, nemohol tušiť, aký revolučný obrat prinesie do medicíny táto koncepcia [1]. Je ohromujúce uvedomiť si, že zhruba tridsať triliónov ľudských buniek a dvadsaťtisíc jeho génov, ktoré považujeme za fyzický základ človeka, ho tvorí len zo 43 % a 1 %; zostatok, teda 39 triliónov buniek a 99 % génov tela človeka patrí mikrobiómu. Tak ako žiadna iná oblasť medicíny, ani diabetológia nemôže napokon nezahrnúť do svojej pozornosti systém, ktorý obsahuje 99 % genetického materiálu tela, je navyše modifikovateľný a priamo reguluje inzulínovú senzitivitu, zápal a metabolizmus.
Do zhruba prelomu tisícročí boli za prevažnú časť chorobnosti a úmrtnosti na zemeguli (podľa Global Burden of Diseases study – GBD) zodpovedné prenosné choroby [2].
Spôsob civilizovania života ľudskej spoločnosti – sanitácia, potrava, vakcinácia a antibiotiká toto globálne bremeno znížili do takej miery, že prestalo byť hlavnou príčinou globálnej úmrtnosti. Táto nadol smerujúca krivka však bola nahradená strmo stúpajúcou krivkou tzv. civilizačných chorôb, nazvaných v GBD akronymom NCD (Non-Communicable Diseases), ktorý odráža historickú zmenu trendu verejného zdravotníctva z prenosných na neprenosné choroby [3]. Medzi NCD patria zložky metabolického syndrómu (MS), ochorenia respiračného traktu, zápalové črevné choroby, autoimunitné a nádorové ochorenia. Táto výmena kriviek je dnes považovaná okrem iného za priamy dôsledok globálnej krízy diverzity mikrobiómu.
V našom článku, ktorého koncept vznikal počas ostatných rokov na diskusných fórach Slovenskej diabetologickej asociácie, sme sa podujali voviesť tému mikrobiómu do ohniska pozornosti diabetológov ako úvod do problematiky. Preto nemožno nezačať od všeobecného.
Fyziológia zdravého mikrobiómu
V nasledujúcom texte, ak nie je uvedené inak, sa užíva termín mikrobióm pre črevný mikrobióm, pretože ten je pre zdravie dominantný.
Črevný „mikrobióm“ je pojem, ktorý – na rozdiel od termínu mikrobiota – obnáša genómy všetkých mikroorganizmov osídľujúcich črevá vrátane baktérií, vírusov, húb a archea, ale aj zložiek ich prostredia (tzv. theatre of activity), v ktorom sú molekuly ich živín, produktov a pôsobkov ľudského tela. Dve zo zhruba 70 fyloskupín tvoria 90 % všetkých črevných mikroorganizmov: Firmicutes a Bacteroidetes. Ďalšie bohato prítomné kmene zahŕňajú Proteobacteria, Actinobacteria a Verrucomicrobia.
Mikrobióm váži až 1,5 kg, obsahuje 10–100 miliárd až trilióny buniek, 1 500 druhov a desiatky miliónov génov. Je to zároveň najbohatší, najrôznorodejší a najvplyvnejší ekosystém ľudského tela.
Kľúčové meradla zdravého mikrobiómu
Alfa-diverzita
Alfa-diverzita číselne vyjadruje štruktúru mikrobiálnej komunity v rámci jednej vzorky – jej bohatosť (počet unikátnych druhov) a vyrovnanosť (rovnomernosť zastúpenia druhov). Patrí k najvalidovanejším metagenomickým markerom gastrointestinálneho a celkového – aj metabolického zdravia. Zníženie alfa-diverzity koreluje so závažnosťou mnohých chorôb, vrátane NCD. Najpoužívanejšie indexy sú počet druhov (operačná taxonomická jednotka/Operational Taxonomic Unit – OTU) a Shannonov index diverzity.
Beta-diverzita
Beta-diverzita meria (ne)podobnosť mikrobiálnych komunít. Pre meranie sa užívajú štatistické metódy ako napríklad Brayovov-Curtisovov index, alebo Jaccard a Jensen-Shannon kvantifikujú tieto odlišnosti pomocou sofistikovaných zjednodušení, ako napríklad metódou UniFrac, vyvinutou americkou mikrobiologičkou Cathrine Lopuzone a jej mentorom Robom Knightom na základe prevedenia fylogenetickej vzdialenosti taxónov do škály 0–1, a tým bol umožnený prevod terrabytov dát zo sekvenácie genómov mikroorganizmov do bodu na grafe. To revolučným spôsobom sprístupnilo interpretáciu analýzy mikrobiomu pre klinickú prax.
Charakteristiky zdravého mikrobiómu
Zdravý mikrobióm je ekosystém bohatý a rôznorodý ako dažďový prales. Oproti tomu dysbióza sa podobá skôr na púšť alebo „opustené mesto bez zelene, kde prežili len potkani a holuby“.

Črevná bariéra a imunitná homeostáza
Intestinálna bariéra je viacvrstvová štruktúra: (1) samotná mikrobiota (2) dvojvrstvový hlien s tzv. „demilitarizovanou zónou“ bez baktérií v blízkosti buniek sliznice, bohatou na IgA a peptidoglykány z Panethových buniek a glykokalyx z pohárikových buniek (3) epitelové bunky spojené v „tight junctions“ a (4) submukózne imunitné, lymfatické, nervové a krvné štruktúry.
V zdravom stave (eubióza) prebieha prísna kompartmentalizácia medzi črevnou mikrobiotou a imunitným systémom. Mikrobióm je zapojený do regulácie diferenciácie T-lymfocytov (Th1/Th2/Th17/Treg), produkcie IgA, sekrécie antimikrobiálnych peptidov a udržania správneho pomeru CD4+/CD8+ a iných subpopulácií. SCFA (hlavne butyrát) priamo aktivujú GPR43-receptory, stimulujú AMPs, tlmia NF-κB a posilňujú tight junction proteíny.
Eubiózu a dobre fungujúcu črevnú bariéru charakterizuje nízky stupeň zápalu, zachyteného napríklad v sérovej hladine lipopolysacharidu, hsCRP alebo zápalových cytokínov.
Ontogenéza mikrobiómu: od pôrodu po dospelý vek
Vývoj mikrobiómu počas života je kľúčový pre pochopenie jeho dlhodobého vplyvu na zdravie. Pred pôrodom je črevná mikrobiota dieťaťa prakticky sterilná (citlivá sekvenácia však určité množstvo mikroorganizmov predsa len zachytí). Prechod pôrodným kanálom je prvou a najdôležitejšou kolonizačnou udalosťou – vaginálna a črevná flóra pôrodných ciest sú mimoriadne dôležitým, ak nie pre zdravie úplne nevyhnutným spôsobom osídlenia novorodenca zdravou mikrobiótou.

Antibiotiká sú najdeštruktívnejším faktorom raného vývoja mikrobiómu. Dieťa v hospodársky rozvinutých krajinách dostane v priemere 3 kúry antibiotík v prvých 2 rokoch života. Podľa rozsiahlych výskumov prof. Blazera – autora knihy Missing Microbes – koreluje vývoj v priebehu desaťročí geografickej distribúcie používania antibiotík v detstve s mapou prevalencie obezity v detstve [4]. Po období stability v dospelosti sa mikrobióm začína opäť meniť a je jedným z mechanizmov starnutia; podľa analýzy črevného mikrobiómu sa dá určiť biologický vek jedinca s mimoriadnou presnosťou a môže sa od chronologického veku odlišovať až o vyše 10 rokov! Teda ak má sedemdesiatnik eubiózu, má biologické charakteristiky a prognózu ako šesťdesiatnik, a naopak – päťdesiatnik s dysbiózou môže mať prognózu ako vyše sedemdesiatročný jedinec.
Dysbióza: patofyziológia
Definícia a mechanizmy
Dysbióza je stav, v ktorom je zloženie a počet črevných baktérií narušený. Ide o poruchu pomeru grampozitívnych a gramnegatívnych baktérií v porovnaní so zdravými jedincami. Klinicky sa manifestuje ako nízka diverzita, nízka abundancia symbiotických druhov a prerastanie patogénnych kmeňov.
Poruchy v zložení črevnej mikrobioty spôsobujú zmeny permeability črevnej bariéry – tzv. leaky gut. Penetrácia endotoxínov (LPS, DAMPs, PAMPs) cez narušenú bariéru vedie k aktivácii inflamasómov – intracelulárnych proteínových komplexov zložených z kaspáz a molekúl derivovaných z fragmentov patogénov a poškodených buniek.
Molekulárne kaskády dysbiózy
LPS a TLR-signalizácia
Lipopolysacharid (LPS) je amfifilný endotoxín v membráne gramnegatívnych baktérií (Bacteroidetes, Proteobacteria). Pri narušení črevnej bariéry LPS vstupuje do portálnej krvi, aktivuje TLR4-receptory na Kupfferových bunkách, hepatocytoch a hviezdicových bunkách pečene, čo spúšťa kaskádu NF-κB a produkciu prozápalových cytokínov (TNFα, IL1β, IL6).
Imunitná dysregulácia
Črevná mikrobiota reguluje špecifické a nešpecifické imunitné mechanizmy stimuláciou lymfoidného tkaniva sliznice (MALT), čo prispieva k sekrécii IgG, IgM, IgA a sIgA. Kľúčová je diferenciácia naivných T-lymfocytov: pomer Th1 (prozápalový)/Th2 (B-lymfocyty protilátky)/Th17 (IL17, regulácia adipogenézy, zápal)/Treg (protizápalový) je priamo modulovaný zložením mikrobioty. IL17, produkovaný Th17 lymfocytmi, je silným regulátorom zápalovej odpovede vrátane vrodenej a adaptívnej imunity, a zároveň regulátorom adipogenézy – čo ho robí relevantným pre diabetológiu.
Hlavné príčiny globálnej krízy diverzity mikrobiómu
Obyvatelia hospodársky rozvinutých krajín sveta žijú uprostred bezprecedentnej krízy mikrobiomu, ktorá bola spôsobená drastickou zmenou životosprávy za ostatných menej ako sto rokov. Zatiaľ čo ľudský genóm zostal na vývojovej úrovni spoločnosti lovcov a zberačov (výskumom mikrobiómu a zdravia u komunít ľudí, žijúcich spôsobom života na úrovni doby kamennej sa venuje rad prác publikovaných v časopise Nature aj prezentácie manželov Sonnenburgovcov na YouTube), mikrobióm „civilizovaného“ človeka sa za posledných 50 rokov zmenil z dažďového pralesa na púšť.
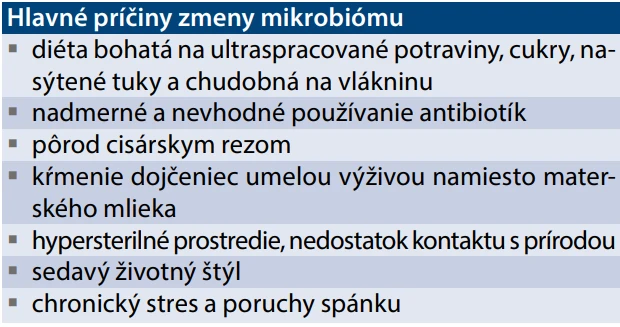
Nedostatok vlákniny a nadmerná konzumácia fruktózy (predovšetkým z tzv. high-fructose corn syrup) spôsobuje dysbiózu posunom v zložení mikrobiómu, zvyšuje črevnú priepustnosť a následne prispieva k patogenéze MS, MASLD a NCD. Nadmerná konzumácia červeného mäsa spôsobuje vysoký prísun L-karnitínu, ktorý je metabolizovaný črevnou mikrobiotou na TMAO (trimetylamín N-oxid) a asociovaný s vyšším rizikom kardiovaskulárnych ochorení a iných NCD. Alkohol znižuje Bacteroidetes, zvyšuje Proteobacteria a Enterobacteriaceae a narúša tight junctions acetaldehydom.
Dysbióza a obezita
Mikrobióm ako regulátor energetickej bilancie
Tzv. obezitogénny mikrobióm má zmenené zloženie, ktoré je asociované so zvýšenou produkciou živín a nadmerným energetickým ziskom z potravy. Prelomová štúdia Turnbaugha et al preukázala, že transplantácia fekálnej mikrobioty z obéznych myší do zárodočne sterilných myší viedla k výraznému nárastu tukového tkaniva bez zmeny príjmu kalórií – tzv. obesity-associated gut microbiome with increased capacity for energy harvest [5].
Elegantným spôsobom sa koncept rozhodujúceho podielu črevného mikrobiómu na patogenéze obezity potvrdil v experimente, v ktorom fekálny mikrobióm získaný od párov jednovaječných dvojčiat diskordantných v obezite preniesol túto chorobu na experimentálne zvieratá chované izokaloricky v sterilnom prostredí [6].
Väčšina štúdií poukazuje na zvýšený pomer Firmicutes/Bacteroidetes u jedincov chorých obezitou, hoci metabolická aktivita mikrobiómu môže byť dôležitejšia ako samotné zloženie. (Ultra)spracované potraviny sú chudobnejšie na mikronutrienty ako potraviny nespracované a určité potravinárske aditíva môžu zvyšovať črevnú permeabilitu a mať prozápalový efekt.
Chronický zápal nízkeho stupňa (low-grade inflammation) navodený dysbiózou a „leaky gut“ je dôležitým faktorom v patogenéze inzulínovej rezistencie (IR) a obezity. Tukové tkanivo pacientov je charakterizované hypertrofickými adipocytmi a nadmernou infiltráciou imunitnými bunkami (hlavne monocytmi), ktoré sú zdrojom prozápalových TNFα a IL6. Tieto cytokíny priamo inhibujú inzulínovú signalizáciu. TLR4-receptor je kľúčovým iniciátorom zápalovej odpovede v makrofágoch.
Sonnenburgom definovaný princíp „hungry microbiome eats you“ hovorí, že pri nedostatku vlákniny začína mikrobióm degradovať mucínový glykokalyx – črevnú obrannú vrstvu [7].

Zdravý mikrobióm je odolný – zdravý mikrobióm dospelých sa vyznačuje tendenciou vrátiť sa k pôvodnému zloženiu po krátkodobých enviromentálnych zmenách, napr. antibiotikách. Zmeny v stravovaní a iných zložkách životosprávy (pohyb, spánok, duševná harmónia) preto môžu mať trvalý efekt na prechod z dysbiózy do eubiózy iba v prípade, že sú trvalé.
Dysbióza a diabetes mellitus 2. typu
Diabetes mellitus 2. typu (DM2T) je spojený so zmenami v zložení črevnej mikroflóry, ktoré mu predchádzajú. Viaceré štúdie našli významné odlišnosti črevného mikrobiómu medzi jedincami zdravými, jedincami s poruchou glukózovej tolerancie a pacientmi s DM2T a aj boli identifikované konzistentné bakteriálne signatúry [8]. U pacientov s DM2T sa pozoruje predovšetkým pokles baktérií Roseburia intestinalis a Faecalibacterium prausnitzii – baktérií zodpovedných za produkciu butyrátu (kľúčového zdroja energie pre kolonocyty a regulátora zápalovej odpovede); a nárast Bacteroides caccae, Clostridiales, Escherichia coli, Desulfovibrio a Lactobacillus spp; následne dochádza k zvýšeniu hladín LPS v sére (metabolická endotoxémia), pravdepodobne v dôsledku zvýšenej translokácie bakteriálnych produktov cez narušenú črevnú bariéru.
Mechanizmy vedúce k DM2T
Rozvoj DM2T môže byť urýchlený komplexnou interakciou medzi imunitným systémom a mikrobiotou vedúcou ku chronickému zápalu. Nižšie sú uvedené centrálne mechanizmy, ktoré sa na rozvoju podieľajú.
Metabolická endotoxémia
Zvýšené hladiny LPS v cirkulácii aktivujú kaskádu TLR4-NF-κB, inhibujú inzulínový receptor substrat-1 (IRS-1) a spúšťajú produkciu TNFα a IL1β – cytokínov priamo interferujúcich s inzulínovým signalizovaním v pečeni, svaloch a tukovom tkanive. Chronická metabolická endotoxémia je teda priamym patogenetickým článkom medzi dysbiózou a inzulínovou rezistenciou.
Pokles butyrátu
Butyrát nielen živí enterocyty, ale priamo potláča NF-κB v imunitných bunkách, zvyšuje expresiu tight junction proteínov (klaudín, okludín, ZO-1), stimuluje sekréciu GLP1 a PYY cez GPR41/43-receptory a podporuje diferenciáciu Treg-lymfocytov. Deficit butyrátu je teda multimodálne prozápalový a prodiabetogénny.
Zmeny v metabolizme žlčových kyselín
Črevná mikrobiota metabolizuje primárne žlčové kyseliny na sekundárne (deoxycholát, litocholát) prostredníctvom bakteriálnych (BSH) enzýmov. Tieto sekundárne žlčové kyseliny aktivujú TGR5-receptor (stimulácia sekrécie GLP1 z L-buniek) a FXR-receptor (regulácia glukoneogenézy, lipogenézy). Dysbiotické posuny v metabolizme žlčových kyselín preto priamo ovplyvňujú glykemickú homeostázu.
Quorum sensing a interakcie medzi baktériami
Quorum sensing – komunikácia baktérií prostredníctvom signálnych molekúl (acylhomoserin laktóny u gramnegatívnych, modifikované oligopeptidy u grampozitívnych baktérií) a je kľúčový pre udržanie eubiózy a kolonizáciu nových prostredí. Narušenie quorum sensingu prispieva k dysbiotickým stavom charakteristickým pre DM2T.
Metformín a mikrobióm
Metformín – historický zlatý štandard farmakoterapie DM2T – má preukázateľný a výrazný efekt na črevný mikrobióm a mnoho jeho priaznivých účinkov nad rámec kontroly diabetu sa uskutočňuje práve týmto mechanizmom (napr. antikancerogénne účinky). Moduluje zloženie mikrobioty (zvyšuje Akkermansia muciniphila), ovplyvňuje metabolizmus žlčových kyselín a zvyšuje produkciu butyrátu. Časť jeho antidiabetického efektu je dnes pripisovaná práve mikrobiotálnym mechanizmom. To otvorilo otázku, či sa pri klinicky nevysvetliteľne variabilnej odpovedi na metformín nejedná o interindividuálne rozdiely v mikrobióme.
Personalizovaná medicína a diabetes mellitus 2. typu
Prelomová štúdia Zeeviho v časopise Cell ukázala, že hlavným determinantom metabolickej odpovede na definované jedlo je mikrobióm jedinca: jedinci v tejto unikátnej štúdii mali personalizovanú postprandiálnu glykemickú odpoveď na rovnaké jedlo, a táto variabilita bola z dominantnej časti v korelácii so zložením mikrobiómu [9]. To môže byť nová oblasť personalizovanej nutričnej stratégie v diabetológii – založenej na vyšetrení mikrobiómu. Nadväzujúce práce predstavili personalizovaný prístup k modelu chudnutia podľa bazálneho mikrobiómu (pred intervenciou) a genetického skóre, ako aj metaanalýzu ukazujúcu vplyv suplementácie Lactobacillus na glykemické a lipidové indexy [10].
Dysbióza a chronické komplikácie diabetes mellitus 2. typu
DM2T, ak nie je optimálne kontrolovaný, je rizikovým faktorom mnohých orgánových komplikácií: chronickej choroby obličiek (CKD), steatotickej choroby pečene (MAFLD/NAFLD), slepoty, ischemickej choroby mozgu (iCMP) koronárnej choroby srdca, choroby periférnych tepien atď.
Iatcu et al zhrnuli v roku 2022 najdôležitejšie poznatky o úlohe mikrobiómu pri chronických komplikáciách DM2T [11]. Zásadným poznatkom je, že metabolické faktory spájajúce dysbiózou s DM2T – chronický zápal nízkej intenzity a oxidatívny stres – sú tie isté, ktoré ovplyvňujú aj vznik a progresiu orgánových komplikácií. Táto zjednocujúca patofyziologická os dáva zmysel konceptu, že modulácia mikrobiómu môže byť sľubnou stratégiou nielen v manažmente samotného DM2T, ale aj jeho komplikácií.
Dysbióza a diabetická nefropatia
Diabetická nefropatia postihuje približne 40 % pacientov s nedostatočne kontrolovaným diabetom a z tých až 20 % môže progredovať do terminálneho štádia CKD s potrebou dialýzy.
Dysbióza hrá preukázateľnú úlohu v jej patogenéze tým, že produkty bakteriálneho metabolizmu ovplyvňujú vznik a progresiu CKD a CKD ďalej zhoršuje črevnú dysbiózou – ide o bludný kruh. Mikrobiotálne zmeny pri CKD sa týkajú predovšetkým poklesu Bifidobacterium, Lactobacillus, Prevotella, Ruminococcaceae, Roseburia a Faecalibacterium (protizápalové, ochranné pre črevnú bariéru) a nárastu Parabacteroides, Enterococcus, Enterobacteriaceae a Klebsiella. Bakteriálna DNA bola detegovaná v krvi 20 % pacientov s pokročilým CKD v rovnakom rode ako v črevách a so zvýšenými biomarkermi zápalu. Tieto nálezy sa stali predmetom štúdia ako terapeutické ciele, čo viedlo k testovaniu intervencií so suplementáciou L. acidophilus + S. thermophilus + B. longum; L. plantarum A7 v sójovom mlieku; kombinácie L. acidophilus + L. casei + B. bifidum a viacerých iných, s priaznivými efektmi na eGFR, HOMA, krvný tlak a iné [11].
Dysbióza a diabetická retinopatia
Dysbióza pri retinopatii je charakterizovaná signifikantným poklesom Bacteroidetes, Actinobacteria a Pasteurellaceae; nárastom Acidaminococcus, Escherichia a Enterobacter. V 12 z 18 analyzovaných rodov bol zaznamenaný pokles u pacientov s diabetickou retinopatiou. Kľúčovým metabolitom je TMAO – pacienti s retinopatiou mali vyššie plazmatické TMAO a prozápalové cytokíny v zrovnaní s pacientmi bez retinopatie, pričom závažnosť korelovala s výškou TMAO. Rekombinantný L. paracasei v experimente znížil retinálnu gliózu, zápal a stratu kapilár; L. rhamnosus u DM1T myší znížil intraokulárny tlak a glykémiu. Klinické štúdie u ľudí zatiaľ chýbajú – čo je dôležitý nezodpovedaný výskumný cieľ.
Podobne by sa dali uviesť špecifické zmeny v mikrobióme u pacientov s DM2T a jeho ostatnými špecifickými komplikáciami. Závažnosť dysbiózy koreluje so závažnosťou týchto komplikácií a obnova eubiózy je asociovaná so zlepšením symptómov a spomalením ich progresie.
Mikrobióm – praktické odporúčania aj pre diabetológov a základné poradenstvo



Táto práca bola podporená Vnútorným grantom SZU, udeleným LS
Zdroje
-
Lederberg J, McCray AT. `Ome Sweet `Omics-A Genealogical Treasury of Words. Scientists Inc 2001; 15(7): 8.
-
[GBD 2019 Diseases and Injuries Collaborators]. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020; 396(10258): 1204–1222. Dostupné z DOI: <https://doi: 10.1016/S0140–6736(20)30925–9>.
-
Bach JF. The effect of infections on susceptibility to autoimmune and allergic diseases. N Engl J Med 2002; 347(12): 911–920. Dostupné z DOI: <http://doi: 10.1056/NEJMra020100>.
-
Blaser MJ. Missing Microbes. Oneworld Publications 2014. ISBN 9781780744414.
-
Turnbaugh PJ, Ley RE, Mahowald MA et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006; 444(7122): 1027–1031. Dostupné z DOI: <http://doi: 10.1038/nature05414>.
-
Ridaura VK, Faith JJ, Rey FE et al. Gut Microbiota from Twins Discordant for Obesity Modulate Metabolism in Mice. Science 2013; 341(1241214). Dostupné z DOI: <http://doi 10.1126/science.1241214>.
-
Sonnenburg ED, Sonnenburg JL. Starving our Microbial Self: The Deleterious Consequences of a Diet Deficient in Microbiota-Accessible Carbohydrates. Cell Metab 2014; 20(5): 779–786. Dostupné z DOI: <http://doi: 10.1016/j.cmet.2014.07.003>.
-
Karlsson FH, Tremaroli V, Nookaew I. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013; 498 : 99–103. Dostupné z DOI: <https://doi.org/10.1038/nature12198>.
-
Zeevi D, Korem T, Zmora N et al. Personalized Nutrition by Prediction of Glycemic Responses. Cell 2015; 163(5): 1079–1094. Dostupné z DOI: <http://doi: 10.1016/j.cell.2015.11.001>.
-
Cuevas-Sierra A, Milagro FI, Guruceaga E et al. A weight-loss model based on baseline microbiota and genetic scores for selection of dietary treatments in overweight and obese population. Clin Nutr 2022; 41(8): 1712–1723. Dostupné z DOI: <https://doi.org/10.1016/j.clnu.2022.06.008>.
-
Iatcu CO, Steen A, Covasa M. Gut Microbiota and Complications of Type-2 Diabetes. Nutrients 2021; 14(1): 166. Dostupné z DOI: <http://doi: 10.3390/nu14010166>.
-
Gilbert J, Knight R, Blakeslee S. Dirt is Good: the advantage of germs for your child’s developing immune system. St. Martin’s Press: New York 2017. ISBN 9781250132604.
Štítky
Diabetologie Endokrinologie Interní lékařstvíČlánek vyšel v časopise
Forum Diabetologicum

2026 Číslo 1
Nejčtenější v tomto čísle
- Moderná diabetológia sa mení na medicínu komplexných biologických súvislostí
- Patogénne dráhy orgánového poškodenia pri metabolickom syndróme
- Klinické postupy manažmentu prevencie vzniku cholelitiázy u vysoko rizikových pacientov liečených receptorovými agonistami GLP1
- Mikrobióm – „kompendium“ pre diabetológov